Ри-бо-ну-кле-и-но-вая кис-ло-та (РНК) - по-ли-мер, мо-но-ме-ра-ми ко-то-рой яв-ля-ют-ся ри-бо-ну-к-лео-ти-ды. Об-ра-зо-ва-ние по-ли-ме-ра про-ис-хо-дит так же, как и у ДНК, за счет фос-фо-ди-эфир-ной связи между остат-ком фос-фор-ной кис-ло-ты и ри-бо-зой.
Мо-но-ме-ры РНК в со-ста-ве нук-лео-ти-дов со-дер-жат пя-ти-уг-ле-род-ный сахар (пен-то-за), фос-фор-ную кис-ло-ту (оста-ток фос-фор-ной кис-ло-ты) и азо-ти-стое ос-но-ва-ние.

Азо-ти-стые ос-но-ва-ния РНК - ура-цил, ци-то-зин, аде-нин и гу-а-нин. Мо-но-са-ха-рид нук-лео-ти-да РНК пред-став-лен ри-бо-зой
РНК - од-но-це-поч-ная мо-ле-ку-ла зна-чи-тель-но мень-ших раз-ме-ров, чем мо-ле-ку-ла ДНК.
Мо-ле-ку-ла РНК со-дер-жит от 75 до 10 000 нук-лео-ти-дов.
РНК-со-дер-жа-щие ви-ру-сы

Мно-гие ви-ру-сы, на-при-мер вирус грип-па, со-дер-жат в ка-че-стве един-ствен-ной нук-ле-и-но-вой кис-ло-ты мо-ле-ку-лу РНК. РНК-со-дер-жа-щих ви-ру-сов, бо-лез-не-твор-ных для че-ло-ве-ка, боль-ше, чем ДНК-со-дер-жа-щих. Они вы-зы-ва-ют по-лио-ми-е-лит, ге-па-тит А, ост-рые про-студ-ные за-бо-ле-ва-ния.
Ар-бо-ви-ру-сы - ви-ру-сы, ко-то-рые пе-ре-но-сят-ся чле-ни-сто-но-ги-ми. Яв-ля-ют-ся воз-бу-ди-те-ля-ми кле-ще-во-го и япон-ско-го эн-це-фа-ли-та, а также жел-той ли-хо-рад-ки.
Рео-ви-ру-сы, ред-кие воз-бу-ди-те-ли ре-спи-ра-тор-ных и ки-шеч-ных за-бо-ле-ва-ний че-ло-ве-ка, стали пред-ме-том осо-бо-го на-уч-но-го ин-те-ре-са из-за того, что их ге-не-ти-че-ский ма-те-ри-ал пред-став-лен в виде двух-це-поч-ной мо-ле-ку-лы РНК.

Также су-ще-ству-ют ре-тро-ви-ру-сы, ко-то-рые вы-зы-ва-ют ряд он-ко-ло-ги-че-ских за-бо-ле-ва-ний.
В за-ви-си-мо-сти от стро-е-ния и вы-пол-ня-е-мой функ-ции раз-ли-ча-ют три ос-нов-ных типа РНК: ри-бо-сом-ную, транс-порт-ную и ин-фор-ма-ци-он-ную (мат-рич-ную).
1. Ин-фор-ма-ци-он-ная РНК
Как по-ка-за-ли ис-сле-до-ва-ния, ин-фор-ма-ци-он-ная РНК со-став-ля-ет 3-5 % от об-ще-го со-дер-жа-ния РНК в клет-ке. Это од-но-це-поч-ная мо-ле-ку-ла, ко-то-рая об-ра-зо-вы-ва-ет-ся в про-цес-се тран-скрип-ции на одной из цепей мо-ле-ку-лы ДНК. Это свя-за-но с тем, что ДНК у ядер-ных ор-га-низ-мов на-хо-дят-ся в ядре, а син-тез белка про-ис-хо-дит на ри-бо-со-мах в ци-то-плаз-ме, по-это-му воз-ник-ла необ-хо-ди-мость в «по-сред-ни-ке». Функ-цию «по-сред-ни-ка» вы-пол-ня-ет мат-рич-ная РНК, она пе-ре-да-ет ин-фор-ма-цию о струк-ту-ре белка из ядра кле-ток, где на-хо-дит-ся ДНК, к ри-бо-со-мам, где эта ин-фор-ма-ция ре-а-ли-зу-ет-ся.

В за-ви-си-мо-сти от объ-е-ма ко-пи-ру-е-мой ин-фор-ма-ции, мо-ле-ку-ла мат-рич-ной РНК может иметь раз-лич-ную длину.
Боль-шин-ство мат-рич-ных РНК су-ще-ству-ют в клет-ке непро-дол-жи-тель-ное время. В бак-те-ри-аль-ных клет-ках су-ще-ство-ва-ние таких РНК опре-де-ля-ет-ся ми-ну-та-ми, а в клет-ках мле-ко-пи-та-ю-щих (в эрит-ро-ци-тах) син-тез ге-мо-гло-би-на (белка) про-дол-жа-ет-ся после утра-ты эрит-ро-ци-та-ми ядра в те-че-ние несколь-ких дней.
2. Ри-бо-сом-ная РНК
Ри-бо-сом-ные РНК со-став-ля-ют 80 % от всех ри-бо-сом, при-сут-ству-ю-щих в клет-ке. Эти РНК син-те-зи-ру-ют-ся в яд-рыш-ке, а в клет-ке они на-хо-дят-ся в ци-то-плаз-ме, где вме-сте с бел-ка-ми об-ра-зу-ют ри-бо-со-мы. На ри-бо-со-мах про-ис-хо-дит син-тез белка. Здесь «код», за-клю-чен-ный в мат-рич-ную РНК, транс-ли-ру-ет-ся в ами-но-кис-лот-ную по-сле-до-ва-тель-ность мо-ле-ку-лы белка.
3. Транс-порт-ная РНК
Транс-порт-ные РНК (см. Рис. 7) об-ра-зу-ют-ся в ядре на ДНК, а затем пе-ре-хо-дят в ци-то-плаз-му.
![]()
На долю таких РНК при-хо-дит-ся около 10 % от об-ще-го со-дер-жа-ния РНК в клет-ке. Они имеют самые ко-рот-кие мо-ле-ку-лы из 80-100 нук-лео-ти-дов.
Транс-порт-ные РНК при-со-еди-ня-ют к себе ами-но-кис-ло-ту и транс-пор-ти-ру-ют ее к месту син-те-за белка, к ри-бо-со-мам.
Все из-вест-ные транс-порт-ные РНК за счет ком-пле-мен-тар-но-го вза-и-мо-дей-ствия между азо-ти-сты-ми ос-но-ва-ни-я-ми об-ра-зо-вы-ва-ют вто-рич-ную струк-ту-ру, по форме на-по-ми-на-ю-щую лист кле-ве-ра. В мо-ле-ку-ле тРНК есть два ак-тив-ных участ-ка - три-плет ан-ти-ко-дон на одном конце и ак-цеп-тор-ный уча-сток, при-со-еди-ня-ю-щий ами-но-кис-ло-ту, на дру-гом.
Каж-дой ами-но-кис-ло-те со-от-вет-ству-ет ком-би-на-ция из трех нук-лео-ти-дов, ко-то-рая носит на-зва-ние три-плет .

Ко-ди-ру-ю-щие ами-но-кис-ло-ты три-пле-ты - ко-до-ны ДНК - пе-ре-да-ют-ся в виде ин-фор-ма-ции три-пле-тов (ко-до-нов) мРНК. У вер-хуш-ки кле-вер-но-го листа тРНК рас-по-ла-га-ет-ся три-плет нук-лео-ти-дов, ко-то-рый ком-пле-мен-та-рен со-от-вет-ству-ю-ще-му ко-до-ну мРНК. Этот три-плет раз-ли-чен для тРНК, пе-ре-но-ся-щих раз-ные ами-но-кис-ло-ты, и ко-ди-ру-ет имен-но ту ами-но-кис-ло-ту, ко-то-рая пе-ре-но-сят-ся дан-ной тРНК. Он по-лу-чил на-зва-ние ан-ти-ко-дон .
Ак-цеп-тор-ный конец яв-ля-ет-ся «по-са-доч-ной пло-щад-кой» для опре-де-лен-ной ами-но-кис-ло-ты.
Таким об-ра-зом, раз-лич-ные типы РНК пред-став-ля-ют собой еди-ную функ-ци-о-наль-ную си-сте-му, на-прав-лен-ную на ре-а-ли-за-цию на-след-ствен-ной ин-фор-ма-ции через син-тез белка.
Кон-цеп-ция РНК мира за-клю-ча-ет-ся в том, что ко-гда-то очень давно мо-ле-ку-ла РНК могла вы-пол-нять функ-цию как мо-ле-ку-лы ДНК, так и бел-ков.
В живых ор-га-низ-мах прак-ти-че-ски все про-цес-сы про-ис-хо-дят бла-го-да-ря фер-мен-там бел-ко-вой при-ро-ды. Белки, од-на-ко, не могут са-мо-реп-ли-ци-ро-вать-ся и син-те-зи-ру-ют-ся в клет-ки на ос-но-ва-нии ин-фор-ма-ции, за-ло-жен-ной в ДНК. Но и удво-е-ние ДНК про-ис-хо-дит толь-ко бла-го-да-ря уча-стию бел-ков и РНК. Сле-до-ва-тель-но, об-ра-зу-ет-ся за-мкну-тый круг, из-за ко-то-ро-го в рам-ках тео-рии воз-ник-но-ве-ния жизни спон-тан-ное воз-ник-но-ве-ние такой слож-ной си-сте-мы ма-ло-ве-ро-ят-но.
В на-ча-ле 1980-х годов в ла-бо-ра-то-рии уче-ных Чека и Ол-т-ме-на (об-ла-да-те-ли но-бе-лев-ской пре-мии по химии) в США была от-кры-та ка-та-ли-ти-че-ская спо-соб-ность РНК. РНК-ка-та-ли-за-то-ры были на-зва-ны ри-бо-зи-ма-ми (
Ока-за-лось, что ак-тив-ный центр ри-бо-сом тоже со-дер-жит боль-шое ко-ли-че-ство ри-бо-сом-ных РНК. Также РНК спо-соб-ны со-зда-вать двой-ную це-поч-ку и са-мо-реп-ли-ци-ро-вать-ся. То есть РНК могли су-ще-ство-вать пол-но-стью ав-то-ном-но, ка-та-ли-зи-руя ме-та-бо-ли-че-ские ре-ак-ции, на-при-мер син-те-за новых ри-бо-ну-кле-а-ти-дов, и са-мо-вос-про-из-во-дясь, со-хра-няя из по-ко-ле-ния в по-ко-ле-ние ка-та-ли-ти-че-ские свой-ства. На-коп-ле-ние слу-чай-ных му-та-ций при-ве-ло к по-яв-ле-нию РНК, ка-та-ли-зи-ру-ю-щих син-тез опре-де-лен-ных бел-ков, яв-ля-ю-щих-ся более эф-фек-тив-ны-ми ка-та-ли-за-то-ра-ми, в связи с чем эти му-та-ции за-креп-ля-лись в ходе есте-ствен-но-го от-бо-ра. Также воз-ник-ли спе-ци-а-ли-зи-ро-ван-ные хра-ни-ли-ща ге-не-ти-че-ской ин-фор-ма-ции - мо-ле-ку-ла ДНК, а РНК стала по-сред-ни-ком между ДНК и бел-ка-ми.
источник конспекта - http://interneturok.ru/ru/school/biology/10-klass/bosnovy-citologii-b/stroenie-i-funktsii-rnk?seconds=0&chapter_id=98
источник видео - http://www.youtube.com/watch?v=d6jd9E1EqhE
источник видео - http://www.youtube.com/watch?v=vSHIuT3eCyA
источник видео - http://www.youtube.com/watch?v=UiuSfDdOs8Q
источник видео - http://www.youtube.com/watch?v=aZd9DZIdt5Y
источник видео - http://www.youtube.com/watch?v=msXWwcK2kqU
источник презенатции - http://prezentacii.com/biologiya/2473-dnk-i-rnk-nukleinovye-kisloty.html
три основных вида РНК: информационная (иРНК), или матричная (мРНК), рибосомная (рРНК), и транспортная (тРНК). Они различаются по величине молекул и функциям. Все типы РНК синтезируются на ДНК при участии ферментов - РНК-полимераз. Информационная РНК составляет 2-3 % всей клеточной РНК, рибосомная - 80-85, транспортная - около 15 %.
иРНК . она считывает наследственную информацию с участка ДНК и в форме скопированной последовательности азотистых оснований переносит ее в рибосомы, где происходит синтез определенного белка. Каждая из молекул иРНК по порядку расположения нуклеотидов и по размеру соответствует гену в ДНК, с которого она была транскрибирована. В среднем иРНК содержит 1500 нуклеотидов (75- 3000). Каждый триплет (три нуклеотида) на иРНК называется кодоном. От кодона зависит, какая аминокислота встанет в данном месте при синтезе белка.
(тРНК) обладает относительно невысокой молекулярной массой порядка 24-29 тыс. Д и содержит в молекуле от 75 до 90 нуклеотидов. До 10 % всех нуклеотидов тРНК приходится на долю минорных оснований, что, по-видимому, защищает ее от действия гидролитических ферментов.Роль тРНК заключается в том, что они переносят аминокислоты к рибосомам и участвуют в процессе синтеза белка. Каждая аминокислота присоединяется к определенной тРНК. Ряд аминокислот обладает более одной тРНК. К настоящему времени обнаружено более 60 тРНК, которые отличаются между собой первичной структурой (последовательностью оснований). Вторичная структура у всех тРНК представлена в виде клеверного листа с двухцепочным стеблем и тремя одноцепочными). На конце одной из цепей находится акцепторный участок - триплет ЦЦА, к аденину которого присоединяется специфическая аминокислота.
(рРНК) . Они содержат 120-3100 нуклеотидов. Рибосомная РНК накапливается в ядре, в ядрышках. В ядрышки из цитоплазмы транспортируются рибосомные белки, и там происходит спонтанное образование субчастиц рибосом путем объединения белков с соответствующими рРНК. Субчастицы рибосомы вместе или врозь транспортируются через поры ядерной мембраны в цитоплазму.Рибосомы представляют собой органеллы величиной 20- 30 нм. Они построены из двух субчастиц разного размера и формы. На определенных стадиях белкового синтеза в клетке происходит разделение рибосом на субчастицы. Рибосомная РНК служит как бы каркасом рибосом и способствует первоначальному связыванию иРНК с рибосомой в процессе биосинтеза белка.
Генетический код- свойственный всем живым организмам способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов.
Свойства: 1) генетический код триплетный (каждая аминокислота кодируется тремя нуклеотидами); 2) неперекрывающийся (соседние триплеты не имеют общих нуклеотидов); 3) вырожденный (за исключением метионина и триптофана все аминокислоты имеют более одного кодона); 4) универсальный (в основном одинаков для всех живых организмов); 5) в кодонах для одной аминокислоты первые два нуклеотида, как правило, одинаковы, а третий варьирует; 6) имеет линейный порядок считывания и характеризуется колине-арностью, т. е. совпадением порядка расположения кодонов в иРНК с порядком расположения аминокислот в синтезирующейся полипептидной цепи.
Различные виды ДНК и РНК - нуклеиновых кислот - это один из объектов изучения молекулярной биологии. Одним из наиболее многообещающих и быстро развивающихся направлений в этой науке в последние годы стало исследование РНК.
Кратко о строении РНК
Итак, РНК, рибонуклеиновая кислота, - это биополимер, молекула которого представляет собой цепочку, образованную четырьмя видами нуклеотидов. Каждый нуклеотид, в свою очередь, состоит из азотистого основания (аденина А, гуанина Г, урацила У либо цитозина Ц) в соединении с сахаром рибозой и остатком фосфорной кислоты. Фосфатные остатки, соединяясь с рибозами соседних нуклеотидов, «сшивают» составные блоки РНК в макромолекулу - полинуклеотид. Так образуется первичная структура РНК.
Вторичная структура - образование двойной цепочки - образуется на некоторых участках молекулы в соответствии с принципом комплементарности азотистых оснований: аденин образует пару с урацилом посредством двойной, а гуанин с цитозином - тройной водородной связи.
В рабочей форме молекула РНК образует также третичную структуру - особое пространственное строение, конформацию.
Синтез РНК
Все виды РНК синтезируются при помощи фермента РНК-полимеразы. Она может быть ДНК- и РНК-зависимой, то есть катализировать синтез как на ДНК, так и на РНК-матрице.
Синтез основан на комплементарности оснований и антипараллельности направления чтения генетического кода и протекает в несколько этапов.
Сначала происходит узнавание и связывание РНК-полимеразы с особой последовательностью нуклеотидов на ДНК - промотором, после чего двойная спираль ДНК раскручивается на небольшом участке и начинается сборка молекулы РНК над одной из цепочек, называемой матричной (другая цепочка ДНК называется кодирующей - именно ее копией является синтезируемая РНК). Асимметричность промотора определяет, какая из цепочек ДНК будет служить матрицей, и тем самым позволяет РНК-полимеразе инициировать синтез в правильном направлении.
Следующий этап называется элонгацией. Транскрипционный комплекс, включающий РНК-полимеразу и расплетенный участок с гибридом ДНК-РНК, начинает движение. По мере этого перемещения наращиваемая цепочка РНК постепенно отделяется, а двойная спираль ДНК расплетается перед комплексом и восстанавливается за ним.

Завершающий этап синтеза наступает, когда РНК-полимераза достигает особого участка матрицы, называемого терминатором. Терминация (окончание) процесса может достигаться различными способами.
Основные виды РНК и их функции в клетке
Они следующие:
- Матричная или информационная (мРНК). Посредством ее осуществляется транскрипция - перенос генетической информации с ДНК.
- Рибосомная (рРНК), обеспечивающая процесс трансляции - синтез белка на матрице мРНК.
- Транспортная (тРНК). Производит узнавание и транспортировку аминокислоты на рибосому, где происходит синтез белка, а также принимает участие в трансляции.
- Малые РНК - обширный класс молекул небольшой длины, осуществляющих разнообразные функции в ходе процессов транскрипции, созревания РНК, трансляции.
- РНК-геномы - кодирующие последовательности, которые содержат генетическую информацию у некоторых вирусов и вироидов.
В 1980-х годах была открыта каталитическая активность РНК. Молекулы, обладающие этим свойством, получили название рибозимов. Естественных рибозимов пока известно не так много, каталитическая способность их ниже, чем у белков, однако в клетке они выполняют исключительно важные функции. В настоящее время ведутся успешные работы по синтезу рибозимов, имеющие в том числе и прикладное значение.
Остановимся подробнее на различных видах молекул РНК.
Матричная (информационная) РНК
Эта молекула синтезируется над расплетенным участком ДНК, копируя таким образом ген, кодирующий тот или иной белок.
РНК эукариотических клеток, прежде чем стать, в свою очередь, матрицей для синтеза белка, должны созреть, то есть пройти через комплекс различных модификаций - процессинг.
Прежде всего, еще на стадии транскрипции, молекула подвергается кэпированию: к ее концу присоединяется особая структура из одного или нескольких модифицированных нуклеотидов - кэп. Он играет важную роль во многих последующих процессах и повышает стабильность мРНК. К другому концу первичного транскрипта присоединяется так называемый поли(А)хвост - последовательность адениновых нуклеотидов.
После этого пре-мРНК подвергается сплайсингу. Это удаление из молекулы некодирующих участков - интронов, которых много в ДНК эукариот. Далее происходит процедура редактирования мРНК, при которой химически модифицируется ее состав, а также метилирование, после чего зрелая мРНК покидает клеточное ядро.

Рибосомная РНК
Основу рибосомы - комплекса, обеспечивающего белковый синтез, составляют две длинные рРНК, которые образуют субчастицы рибосомы. Синтезируются они совместно в виде одной пре-рРНК, которая затем в ходе процессинга разделяется. В большую субчастицу входит также низкомолекулярная рРНК, синтезируемая с отдельного гена. Рибосомные РНК обладают плотно упакованной третичной структурой, которая служит каркасом для белков, присутствующих в рибосоме и выполняющих вспомогательные функции.
В нерабочей фазе субъединицы рибосомы разделены; при инициации трансляционного процесса рРНК малой субчастицы соединяется с матричной РНК, после чего происходит полное объединение элементов рибосомы. При взаимодействии РНК малой субчастицы с мРНК последняя как бы протягивается через рибосому (что равнозначно движению рибосомы по мРНК). Рибосомная РНК большой субчастицы является рибозимом, то есть обладает ферментными свойствами. Она катализирует образование пептидных связей между аминокислотами в ходе синтеза белка.

Следует отметить, что наибольшая часть всей РНК в клетке приходится на долю рибосомной - 70-80 %. ДНК обладает большим количеством генов, кодирующих рРНК, что обеспечивает весьма интенсивную ее транскрипцию.
Транспортная РНК
Эта молекула распознается определенной аминокислотой при помощи особого фермента и, соединяясь с ней, производит транспортировку аминокислоты на рибосому, где служит посредником в процессе трансляции - синтеза белка. Перенос осуществляется путем диффузии в цитоплазме клетки.
Вновь синтезированные молекулы тРНК, так же как и другие виды РНК, подвергаются процессингу. Зрелая тРНК в активной форме имеет конформацию, напоминающую клеверный лист. На «черешке» листа - акцепторном участке - расположена последовательность ЦЦА с гидроксильной группой, которая связывается с аминокислотой. На противоположном конце «листа» находится антикодоновая петля, которая соединяется с комплементарным кодоном на мРНК. D-петля служит для связывания транспортной РНК с ферментом при взаимодействии с аминокислотой, а Т-петля - для связывания с большой субчастицей рибосомы.

Малые РНК
Эти виды РНК играют важную роль в клеточных процессах и сейчас активно изучаются.
Так, например, малые ядерные РНК в клетках эукариот участвуют в сплайсинге мРНК и, возможно, обладают каталитическими свойствами наряду с белками сплайсосом. Малые ядрышковые РНК участвуют в процессинге рибосомной и транспортной РНК.
Малые интерферирующие и микроРНК являются важнейшими элементами системы регуляции экспрессии генов, необходимой клетке для контроля собственной структуры и жизнедеятельности. Эта система - важная часть иммунного антивирусного ответа клетки.
Существует также класс малых РНК, функционирующих в комплексе с белками Piwi. Эти комплексы играют огромную роль в развитии клеток зародышевой линии, в сперматогенезе и в подавлении мобильных генетических элементов.
РНК-геном
Молекула РНК может использоваться в качестве генома большинством вирусов. Вирусные геномы бывают различными - одно- и двухцепочечными, кольцевыми или линейными. Также РНК-геномы вирусов часто бывают сегментированы и в целом короче, чем ДНК-содержащие геномы.
Существует семейство вирусов, генетическая информация которых, закодированная в РНК, после инфицирования клетки путем обратной транскрипции переписывается на ДНК, которая затем внедряется в геном клетки-жертвы. Это так называемые ретровирусы. К ним, в частности, относится вирус иммунодефицита человека.

Значение исследования РНК в современной науке
Если прежде преобладало мнение о второстепенной роли РНК, то ныне ясно, что она - необходимый и важнейший элемент внутриклеточной жизнедеятельности. Множество процессов первостепенной значимости не обходятся без активного участия РНК. Механизмы таких процессов долгое время оставались неизвестными, но благодаря исследованию различных видов РНК и их функций постепенно проясняются многие детали.
Не исключено, что РНК сыграла решающую роль в возникновении и становлении жизни на заре истории Земли. Результаты недавних исследований говорят в пользу этой гипотезы, свидетельствуя о необычайной древности многих механизмов функционирования клетки с участием тех или иных видов РНК. Например, недавно открытые рибопереключатели в составе мРНК (система безбелковой регуляции активности генов на стадии транскрипции), по мнению многих исследователей, являются отголосками эпохи, когда примитивная жизнь строилась на основе РНК, без участия ДНК и белков. Также весьма древним компонентом системы регуляции считаются микроРНК. Особенности структуры каталитически активной рРНК свидетельствуют о ее постепенной эволюции путем присоединения новых фрагментов к древней проторибосоме.
Тщательное изучение того, какие виды РНК и каким образом заняты в тех или иных процессах, исключительно важно также для теоретических и прикладных областей медицины.
К нуклеиновым кислотам относят высокополимерные соединения, распадающиеся при гидролизе на пуриновые и пиримидиновые основания, пентозу и фосфорную кислоту. Нуклеиновые кислоты содержат углерод, водород, фосфор, кислород и азот. Различают два класса нуклеиновых кислот: рибонуклеиновые кислоты (РНК) и дезоксирибонуклеиновые кислоты (ДНК) .
Строение и функции ДНК
ДНК — полимер, мономерами которой являются дезоксирибонуклеотиды. Модель пространственного строения молекулы ДНК в виде двойной спирали была предложена в 1953 г. Дж. Уотсоном и Ф. Криком (для построения этой модели они использовали работы М. Уилкинса, Р. Франклин, Э. Чаргаффа).
Молекула ДНК образована двумя полинуклеотидными цепями, спирально закрученными друг около друга и вместе вокруг воображаемой оси, т.е. представляет собой двойную спираль (исключение — некоторые ДНК-содержащие вирусы имеют одноцепочечную ДНК). Диаметр двойной спирали ДНК — 2 нм, расстояние между соседними нуклеотидами — 0,34 нм, на один оборот спирали приходится 10 пар нуклеотидов. Длина молекулы может достигать нескольких сантиметров. Молекулярный вес — десятки и сотни миллионов. Суммарная длина ДНК ядра клетки человека — около 2 м. В эукариотических клетках ДНК образует комплексы с белками и имеет специфическую пространственную конформацию.
Мономер ДНК — нуклеотид (дезоксирибонуклеотид) — состоит из остатков трех веществ: 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) и 3) фосфорной кислоты. Азотистые основания нуклеиновых кислот относятся к классам пиримидинов и пуринов. Пиримидиновые основания ДНК (имеют в составе своей молекулы одно кольцо) — тимин, цитозин. Пуриновые основания (имеют два кольца) — аденин и гуанин.
Моносахарид нуклеотида ДНК представлен дезоксирибозой.
Название нуклеотида является производным от названия соответствующего основания. Нуклеотиды и азотистые основания обозначаются заглавными буквами.
Полинуклеотидная цепь образуется в результате реакций конденсации нуклеотидов. При этом между 3"-углеродом остатка дезоксирибозы одного нуклеотида и остатком фосфорной кислоты другого возникает фосфоэфирная связь (относится к категории прочных ковалентных связей). Один конец полинуклеотидной цепи заканчивается 5"-углеродом (его называют 5"-концом), другой — 3"-углеродом (3"-концом).
Против одной цепи нуклеотидов располагается вторая цепь. Расположение нуклеотидов в этих двух цепях не случайное, а строго определенное: против аденина одной цепи в другой цепи всегда располагается тимин, а против гуанина — всегда цитозин, между аденином и тимином возникают две водородные связи, между гуанином и цитозином — три водородные связи. Закономерность, согласно которой нуклеотиды разных цепей ДНК строго упорядоченно располагаются (аденин — тимин, гуанин — цитозин) и избирательно соединяются друг с другом, называется принципом комплементарности . Следует отметить, что Дж. Уотсон и Ф. Крик пришли к пониманию принципа комплементарности после ознакомления с работами Э. Чаргаффа. Э. Чаргафф, изучив огромное количество образцов тканей и органов различных организмов, установил, что в любом фрагменте ДНК содержание остатков гуанина всегда точно соответствует содержанию цитозина, а аденина — тимину («правило Чаргаффа» ), но объяснить этот факт он не смог.
Из принципа комплементарности следует, что последовательность нуклеотидов одной цепи определяет последовательность нуклеотидов другой.
Цепи ДНК антипараллельны (разнонаправлены), т.е. нуклеотиды разных цепей располагаются в противоположных направлениях, и, следовательно, напротив 3"-конца одной цепи находится 5"-конец другой. Молекулу ДНК иногда сравнивают с винтовой лестницей. «Перила» этой лестницы — сахарофосфатный остов (чередующиеся остатки дезоксирибозы и фосфорной кислоты); «ступени» — комплементарные азотистые основания.
Функция ДНК — хранение и передача наследственной информации.
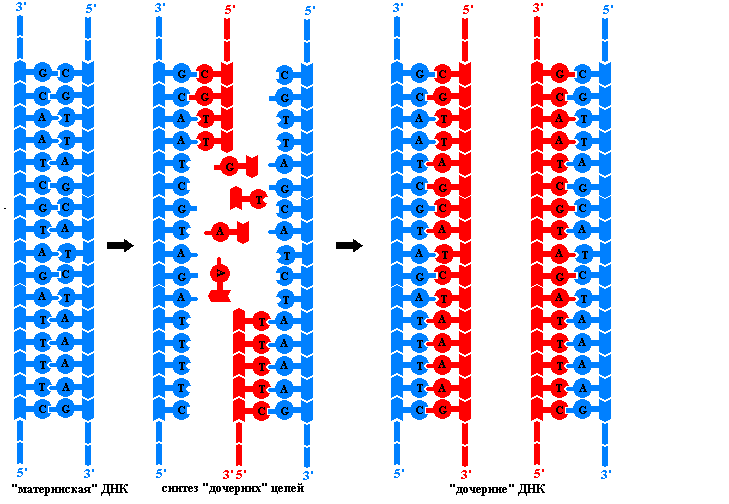
Репликация (редупликация) ДНК
— процесс самоудвоения, главное свойство молекулы ДНК. Репликация относится к категории реакций матричного синтеза, идет с участием ферментов. Под действием ферментов молекула ДНК раскручивается, и около каждой цепи, выступающей в роли матрицы, по принципам комплементарности и антипараллельности достраивается новая цепь. Таким образом, в каждой дочерней ДНК одна цепь является материнской, а вторая — вновь синтезированной. Такой способ синтеза называется полуконсервативным .
«Строительным материалом» и источником энергии для репликации являются дезоксирибонуклеозидтрифосфаты (АТФ, ТТФ, ГТФ, ЦТФ), содержащие три остатка фосфорной кислоты. При включении дезоксирибонуклеозидтрифосфатов в полинуклеотидную цепь два концевых остатка фосфорной кислоты отщепляются, и освободившаяся энергия используется на образование фосфодиэфирной связи между нуклеотидами.

В репликации участвуют следующие ферменты:
- геликазы («расплетают» ДНК);
- дестабилизирующие белки;
- ДНК-топоизомеразы (разрезают ДНК);
- ДНК-полимеразы (подбирают дезоксирибонуклеозидтрифосфаты и комплементарно присоединяют их к матричной цепи ДНК);
- РНК-праймазы (образуют РНК-затравки, праймеры);
- ДНК-лигазы (сшивают фрагменты ДНК).
С помощью геликаз в определенных участках ДНК расплетается, одноцепочечные участки ДНК связываются дестабилизирующими белками, образуется репликационная вилка . При расхождении 10 пар нуклеотидов (один виток спирали) молекула ДНК должна совершить полный оборот вокруг своей оси. Чтобы предотвратить это вращение ДНК-топоизомераза разрезает одну цепь ДНК, что дает ей возможность вращаться вокруг второй цепи.
ДНК-полимераза может присоединять нуклеотид только к 3"-углероду дезоксирибозы предыдущего нуклеотида, поэтому данный фермент способен передвигаться по матричной ДНК только в одном направлении: от 3"-конца к 5"-концу этой матричной ДНК. Так как в материнской ДНК цепи антипараллельны, то на ее разных цепях сборка дочерних полинуклеотидных цепей происходит по-разному и в противоположных направлениях. На цепи 3"-5" синтез дочерней полинуклеотидной цепи идет без перерывов; эта дочерняя цепь будет называться лидирующей . На цепи 5"-3" — прерывисто, фрагментами (фрагменты Оказаки ), которые после завершения репликации ДНК-лигазами сшиваются в одну цепь; эта дочерняя цепь будет называться запаздывающей (отстающей ).
Особенностью ДНК-полимеразы является то, что она может начинать свою работу только с «затравки» (праймера ). Роль «затравок» выполняют короткие последовательности РНК, образуемые при участи фермента РНК-праймазы и спаренные с матричной ДНК. РНК-затравки после окончания сборки полинуклеотидных цепочек удаляются.
Репликация протекает сходно у прокариот и эукариот. Скорость синтеза ДНК у прокариот на порядок выше (1000 нуклеотидов в секунду), чем у эукариот (100 нуклеотидов в секунду). Репликация начинается одновременно в нескольких участках молекулы ДНК. Фрагмент ДНК от одной точки начала репликации до другой образует единицу репликации — репликон .
Репликация происходит перед делением клетки. Благодаря этой способности ДНК осуществляется передача наследственной информации от материнской клетки дочерним.
Репарация («ремонт»)
Репарацией называется процесс устранения повреждений нуклеотидной последовательности ДНК. Осуществляется особыми ферментными системами клетки (ферменты репарации ). В процессе восстановления структуры ДНК можно выделить следующие этапы: 1) ДНК-репарирующие нуклеазы распознают и удаляют поврежденный участок, в результате чего в цепи ДНК образуется брешь; 2) ДНК-полимераза заполняет эту брешь, копируя информацию со второй («хорошей») цепи; 3) ДНК-лигаза «сшивает» нуклеотиды, завершая репарацию.
Наиболее изучены три механизма репарации: 1) фоторепарация, 2) эксцизная, или дорепликативная, репарация, 3) пострепликативная репарация.
Изменения структуры ДНК происходят в клетке постоянно под действием реакционно-способных метаболитов, ультрафиолетового излучения, тяжелых металлов и их солей и др. Поэтому дефекты систем репарации повышают скорость мутационных процессов, являются причиной наследственных заболеваний (пигментная ксеродерма, прогерия и др.).
Строение и функции РНК

— полимер, мономерами которой являются рибонуклеотиды . В отличие от ДНК, РНК образована не двумя, а одной полинуклеотидной цепочкой (исключение — некоторые РНК-содержащие вирусы имеют двухцепочечную РНК). Нуклеотиды РНК способны образовывать водородные связи между собой. Цепи РНК значительно короче цепей ДНК.
Мономер РНК — нуклеотид (рибонуклеотид) — состоит из остатков трех веществ: 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) и 3) фосфорной кислоты. Азотистые основания РНК также относятся к классам пиримидинов и пуринов.
Пиримидиновые основания РНК — урацил, цитозин, пуриновые основания — аденин и гуанин. Моносахарид нуклеотида РНК представлен рибозой.
Выделяют три вида РНК : 1) информационная (матричная) РНК — иРНК (мРНК), 2) транспортная РНК — тРНК, 3) рибосомная РНК — рРНК.
Все виды РНК представляют собой неразветвленные полинуклеотиды, имеют специфическую пространственную конформацию и принимают участие в процессах синтеза белка. Информация о строении всех видов РНК хранится в ДНК. Процесс синтеза РНК на матрице ДНК называется транскрипцией.

Транспортные РНК содержат обычно 76 (от 75 до 95) нуклеотидов; молекулярная масса — 25 000-30 000. На долю тРНК приходится около 10% от общего содержания РНК в клетке. Функции тРНК: 1) транспорт аминокислот к месту синтеза белка, к рибосомам, 2) трансляционный посредник. В клетке встречается около 40 видов тРНК, каждый из них имеет характерную только для него последовательность нуклеотидов. Однако у всех тРНК имеется несколько внутримолекулярных комплементарных участков, из-за которых тРНК приобретают конформацию, напоминающую по форме лист клевера. У любой тРНК есть петля для контакта с рибосомой (1), антикодоновая петля (2), петля для контакта с ферментом (3), акцепторный стебель (4), антикодон (5). Аминокислота присоединяется к 3"-концу акцепторного стебля. Антикодон — три нуклеотида, «опознающие» кодон иРНК. Следует подчеркнуть, что конкретная тРНК может транспортировать строго определенную аминокислоту, соответствующую ее антикодону. Специфичность соединения аминокислоты и тРНК достигается благодаря свойствам фермента аминоацил-тРНК-синтетаза.
Рибосомные РНК содержат 3000-5000 нуклеотидов; молекулярная масса — 1 000 000-1 500 000. На долю рРНК приходится 80-85% от общего содержания РНК в клетке. В комплексе с рибосомными белками рРНК образует рибосомы — органоиды, осуществляющие синтез белка. В эукариотических клетках синтез рРНК происходит в ядрышках. Функции рРНК : 1) необходимый структурный компонент рибосом и, таким образом, обеспечение функционирования рибосом; 2) обеспечение взаимодействия рибосомы и тРНК; 3) первоначальное связывание рибосомы и кодона-инициатора иРНК и определение рамки считывания, 4) формирование активного центра рибосомы.
Информационные РНК разнообразны по содержанию нуклеотидов и молекулярной массе (от 50 000 до 4 000 000). На долю иРНК приходится до 5% от общего содержания РНК в клетке. Функции иРНК : 1) перенос генетической информации от ДНК к рибосомам, 2) матрица для синтеза молекулы белка, 3) определение аминокислотной последовательности первичной структуры белковой молекулы.
Строение и функции АТФ
Аденозинтрифосфорная кислота (АТФ) — универсальный источник и основной аккумулятор энергии в живых клетках. АТФ содержится во всех клетках растений и животных. Количество АТФ в среднем составляет 0,04% (от сырой массы клетки), наибольшее количество АТФ (0,2-0,5%) содержится в скелетных мышцах.
АТФ состоит из остатков: 1) азотистого основания (аденина), 2) моносахарида (рибозы), 3) трех фосфорных кислот. Поскольку АТФ содержит не один, а три остатка фосфорной кислоты, она относится к рибонуклеозидтрифосфатам.
Для большинства видов работ, происходящих в клетках, используется энергия гидролиза АТФ. При этом при отщеплении концевого остатка фосфорной кислоты АТФ переходит в АДФ (аденозиндифосфорную кислоту), при отщеплении второго остатка фосфорной кислоты — в АМФ (аденозинмонофосфорную кислоту). Выход свободной энергии при отщеплении как концевого, так и второго остатков фосфорной кислоты составляет по 30,6 кДж. Отщепление третьей фосфатной группы сопровождается выделением только 13,8 кДж. Связи между концевым и вторым, вторым и первым остатками фосфорной кислоты называются макроэргическими (высокоэнергетическими).
Запасы АТФ постоянно пополняются. В клетках всех организмов синтез АТФ происходит в процессе фосфорилирования, т.е. присоединения фосфорной кислоты к АДФ. Фосфорилирование происходит с разной интенсивностью при дыхании (митохондрии), гликолизе (цитоплазма), фотосинтезе (хлоропласты).
АТФ является основным связующим звеном между процессами, сопровождающимися выделением и накоплением энергии, и процессами, протекающими с затратами энергии. Кроме этого, АТФ наряду с другими рибонуклеозидтрифосфатами (ГТФ, ЦТФ, УТФ) является субстратом для синтеза РНК.
Перейти к лекции №3 «Строение и функции белков. Ферменты»
Перейти к лекции №5 «Клеточная теория. Типы клеточной организации»
Молекула РНК также полимер, мономерами которого является рибонуклеотиды, РНК представляет собой одноцепочную молекулу. Она построена таким же образом, как и одна из цепей ДНК. Нуклеотиды РНК похожи на нуклеотиды ДНК, хотя и не тождественны им. Их тоже четыре, и они состоят из осатков азотистого основания, пентозы и фосфорной кислоты. Три азотистых основания совершенно такие же, как в ДНК: А , Г и Ц . Однако вместо Т у ДНК в РНК присутствует близкое по строение пиримидиновое основание – урацил (У ). основное различие между ДНК и РНК – это характер углевода: в нуклотидах ДНК моносахарид – дезоксирибоза, а в РНК – рибоза. Связь между нуклеотидами осуществляется, как и в ДНК, через сахар и остаток фосфорной кислоты. В отличие от ДНК, содержание которой в клетках определенных организмов постоянно, содержание РНК в них колеблется. Оно заметно выше там, где происходит интенсивный синтез.
В отношении выполняемых функций различают несколько видов РНК.
Транспортная РНК (тРНК). Молекулы тРНк самые короткие: они состоят всего из 80-100 нуклеотидов. Молекулярная масса таких частиц равна 25-30 тыс. Транспортные РНК в основном содержатся в цитоплазме клетки. Функция их состоит в переносе аминокислот в рибосомы, к месту синтеза белка. Из общего содержания РНК клетк на долю тРНК приходится около 10%.
Рибосомная РНК (рРНК). Это крупные молекулы: в их состав входит 3-5 тыс. нуклеотидов, соотвественно их молекулярная масса достигает 1-1,5 млн. Рибосомные РНК составляют существенную часть рибосомы. Из общего содержания РНК в клетке на долю рРНК приходится около 90%.
Информационная РНК (иРНК), или матричная РНК (мРНК), содержится в ядре и цитоплазме. Функция ее состоит в переносе информации о структуре белка от ДНК к месту синтеза белка в рибосомах. На долю иРНК приходится примерно 0,5-1% от общего содержания РНК клетки. Размер иРНК колеблется в широких пределах – от 100 до 10000 нуклеотидов.
Все виды РНК синтезируются на ДНК, которая служитсвоего рода матрицей.
ДНК – носитель наследсвенной нформации.
Каждый белок представлен одной или несколькими полипиптидными цепями. Участок ДНК, несущий информацию об одной полипиптидной цепи, называют геном . Совокупность молекул ДНК клетки выполняет функцию носителя генетической информации. Генетическая информация передается как от материнской клетки дочерним клеткам, так и от родителей детям. Ген является единицей генетической , или наследственной, информации.
ДНК – носитель генетической информаци в клетке – непосредственного участия в синтезе белков не принимает. В клетках эукариот молекулы ДНК содержатся в хромосомах ядра и отделены ядерной оболочкой от цитоплазмы, где происходит синтез белков. К рибосомам – местам сборки белков – высылается из ядра несущий информацию посредник, способный пройти через поры ядерной оболочки. Таким посредником является информационная РНК (иРНК). По принципу комплементарности она синтезируется на ДНК при участие фермента, называемого РНК-полимеразой .

Информационная РНК – это однонитевая молекула, и транскрипция идет с одной цепи двунитевой молекулы ДНК. Она является копией не всей молекулы ДНК, а только части ее – одного гена у эукариот или группы рядом расположенных генов, несущих информацию о структуре белков, необходимых для выполнения одной функции, у прокариот. Такую группу генов называют опероном . В начале каждого оперона находится своего рода посадочная площадка для РНК-полимеразы, называемая промотором .это специфическая последовательность нуклеотидов ДНК, которую фермент «узнает» благодаря химическому сродству. Только присоединившись к промотору, РНК-полиммераза способна начать интез РНК. Доядя до конца оперона, фермент встречает сигнал (в виде определенной последоватльности нуклеотидов), означающий конец считывания. Готовая иРНК отходит от ДНК и направляется к месту синтеза белков.
В процессе транскрипции можно выделить четыре стадии: 1) связывание РНК -полимеразы с промотором; 2) инициация – начало синтеза. Оназаключается в образовании первой фосфодиэфирной связи между АТФ или ГТФ и вторым нуклеотидом синтезирующейся молекулы РНК; 3) элонгация – рост цепи РНК; т.е. последовательное присоединение нуклеотидов друг к другу в том порядке, в котором стоят комплементарные им нуклеотиды в транскрибируемой нити ДНК. Скорость элонгации 50 нуклеотидов в секунду; 4) терминация – завершение синтеза РНК.
Пройдя через поры ядерной оболочки, иРНК направляется к рибосомам, где осуществляется расшифровка генетической информации – перевод ее с «языка» нуклеотидов на «язык» аминокислот. Синтез полипептидных цепей по матрице иРНК, происходящий в рибосомах, называют трансляцией (лат. translation – перевод).
Аминокислоты, из котрых синтезируютсябелки, доставляются к рибосомам с помощью специальных РНК, называемых транспортными (тРНК). В клетке имеется столько же разных тРНК, сколько кодонов, шифрующих аминокислоты. На вершине «листа» каждой тРНК имеется последовательность трех нуклеотидов, комплементарных нуклеотидам кодона в иРНК. Ее называют антикодоном. Специальный фермент – кодаза – опознает тРНК и присоединяет к «черешку листа» аминокислоту – только ту, которая кодируется триплетом, комплементарным антикодону. На образование ковалентной связи между тРНК и «своей» аминокислотой затрачивается энергия одной молекулы АТФ.
 Для
того чтобы аминокислота включилась в
полипептидную цепь, она должна оторваться
от тРНК. Это становится возможным, когда
тРНК поступает на рибососму и антикодон
узнает свой кодон в иРНК. В рибосоме
имеется два участка для связывания двух
молекул тРНК. В один из этих участков,
называемый акцепторным
,
поступает тРНК с аминокислотой и
присоединяется к своему кодону (I).
Эта аминокислота присоединяет к себе
(акцептирует) растущую цепь белка (II)?
Между ними образуется пептидная связь.
тРНК, к которой теперь присоединяется
вместе с кодоном иРНК в донорный
участок рибосомы. В освободившийся
акцепторный участок приходит новая
тРНК, связанная с аминокислотой, которая
шифруется очередным кодоном (III).
Из донорного участка сюда вновь
переносится оторвавшаяся полипептидная
цепь и удлинняется еще на одно звено.
Аминокислоты в растущей цепи соединены
в той последовательности, в которой
расположены шифрующие их кодоны в иРНК.
Для
того чтобы аминокислота включилась в
полипептидную цепь, она должна оторваться
от тРНК. Это становится возможным, когда
тРНК поступает на рибососму и антикодон
узнает свой кодон в иРНК. В рибосоме
имеется два участка для связывания двух
молекул тРНК. В один из этих участков,
называемый акцепторным
,
поступает тРНК с аминокислотой и
присоединяется к своему кодону (I).
Эта аминокислота присоединяет к себе
(акцептирует) растущую цепь белка (II)?
Между ними образуется пептидная связь.
тРНК, к которой теперь присоединяется
вместе с кодоном иРНК в донорный
участок рибосомы. В освободившийся
акцепторный участок приходит новая
тРНК, связанная с аминокислотой, которая
шифруется очередным кодоном (III).
Из донорного участка сюда вновь
переносится оторвавшаяся полипептидная
цепь и удлинняется еще на одно звено.
Аминокислоты в растущей цепи соединены
в той последовательности, в которой
расположены шифрующие их кодоны в иРНК.
Когда на рибосоме оказывается один из трех триплетов (УАА, УАГ, УГА ), являющиеся «знаками препинания» между генами, ни одна тРНК не может занять место в акцепторном участке. Дело в том, что не существует антикодонов, комплементарных последовательностям нуклеотидов «знаков препинания». Оторвавшейся цепи не к чему присоединиться в акцепторном участке, и она покидает рибосому. Синтез белка завершен.
У прокариот синтез белков начинается с того, что кодон АУГ , расположенный на первом месте в копии с каждого гена, занимае в рибосоме такую позицию, что с ним взаимодействует антикодон особой тРНК, оединенной с формилментионином . Эта измененная форма аминокислоты метионина сразу попадает в донорный участок и выполняет роль заглавной буквы во фразе – с нее в бактериальной клетке начинается синтез любой полипептидной цепи. Когда триплет АУГ стоит не на первом месте, а внутри копии с гена, он кодирует аминокислоту метионин. После завершения синтеза полипептидной цепи формилметионин отщепляется от нее и в готовом белке отсуствует.
 Для
увеличения производства белков иРНК
часто проходит одновременно не по одной,
а по нескольким рибосомам. Акую структуру,
объединенную одной молекулой иРНК,
называют полисомой
.
На каждой рибосоме вэтом похожем на
нитку бус конвейере синтезируются
одинаковые белки.
Для
увеличения производства белков иРНК
часто проходит одновременно не по одной,
а по нескольким рибосомам. Акую структуру,
объединенную одной молекулой иРНК,
называют полисомой
.
На каждой рибосоме вэтом похожем на
нитку бус конвейере синтезируются
одинаковые белки.
Аминокислоты бесперебойно поставляются к рибосомам с помощью тРНК. Отдав аминокислоту, тРНК покидает рибосому и с помощью кодазы соединяется. Высокая слаженность всех «служб комбината» по производсву белов позволяет в течении нескольких секунд синтезировать полипептидные цепи, состоящие из сотен аминокислот.
Свойства генетического кода. Благодаря процессу транскрипции в клетке осуществляется передача информации от ДНК к белку
ДНК → иРНК → белок
Генетическая информация, содержащаяся в ДНК и в иРНК, заключена в последовательности расположения нуклеотидов в молекулах.
 Каким
же образом происходит перевод информации
с «языка» нуклеотидов на «язык»
аминокислот? Такой перевод осуществляется
с помощью генетического кода. Код,
или шифр
,
- это система символов для перевода
одной формы информации в другую.
Генетический
код
–это
система записи информации о
последовательности расположения
аминокислот в белках с помощью
последовательности расположения
нуклеотидов в иРНК.
Каким
же образом происходит перевод информации
с «языка» нуклеотидов на «язык»
аминокислот? Такой перевод осуществляется
с помощью генетического кода. Код,
или шифр
,
- это система символов для перевода
одной формы информации в другую.
Генетический
код
–это
система записи информации о
последовательности расположения
аминокислот в белках с помощью
последовательности расположения
нуклеотидов в иРНК.
Какими же свойствами обладает генетический код?
Код триплетен . В состав РНК входят четыре нуклеотида: А, Г, Ц, У. Если бы мы пытались обозначить одну аминокислоту одним нуклеотидом, то 16 из 20 аминокислот остались бы не зашифрованы. Двух буквенный код позволил бы зашифровать 16 аминокислот. Природа создала трехбуквенный, или триплетный, код. Это означает, что каждая из 20 аминокислот зашифрована последовательностью из трех нуклеотидов, называемой триплетом или кодоном.
Код вырожден. Это означает, что каждая аминокислота шифруется более чем одним кодоном. Исключения: метеонин и триптофан, каждая из которых кодируется одним триплетом.
Код однозначен. Каждый кодон шифрует только одну аминокислоту.
Между генами имеется «знаки препинания». В печатном тексте в конце каждой фразы стоит точка. Несколько связанных по смыслу фраз составляют абзац. На языке генетической информации таким абзацем являетсяоперон и комплементарная ему иРНК. Каждый ген в опероне прокариот или отдельный ген эукариот кодирует одну полипептидную цепочку – фразу. Так как в ряде случаев по матрице иРНК последовательно создается несколько разных полипептидных цепей, они должны быть отделены друг от друга. Для этого в генетическом годе имеются три специальных триплета – УАА, УАГ, УГА, каждый из которых обозначает прекращение синтеза одной полипептидной цепи. Таким образом, эти триплеты выполняют функцию знаков препинания. Они находятся в конце каждого гена.
Внутри гена нет «знаков препинания».
Код универсален. Генетический код един для всех живущих на Земле существ. У бактерий и грибов, пшеницы и хлопка, рыб и червей, лягушек и человека одни и те же триплеты кодируют одни и те же аминокислоты.

Принципы репликации ДНК. Преемственность генетического материала в поколениях клеток и организмов обеспечивается процессом репликации – удвоения молекул ДНК. Этот сложный процесс осуществляется комплексом нескольких ферментов и не обладающих каталитической активностью белов, необходимых для придания полинуклеотидным цепям нужной конформации. В результате репликации образуются две идентичные двойные спирали ДНК. Эти так называемые дочерние молекулы ничем не отличаются друг от друга и от исходной материнской молекулы ДНК. Репликация происходит в клетке перед делением, поэтому каждая дочерняя клетка получает точно такие же молекулы ДНК, какие имела материнская клетка. Процесс репликации основан на ряде принципов:
Только
в этом случае ДНК-полимеразы способна
двигаться по материнским нитям и
использовать их в качестве матриц для
безошибочного синтеза дочерних цепей.
Но полное раскручивание спиралей,
состоящих из многих миллионов пар
нуклеотидов, сопряжено со столь
значительным числом вращений и такими
энергетическими затратами, которые
невозможны в условиях клетки. Поэтому
репликация у эукариот начинается
одновременно в некоторых местах молекулы
ДНК. Участок между двумя точками, в
которых начинается синтез дочерних
цепей, называют репликоном
.
Он является единицей
репликации.


В каждой молекуле ДНК эукариотической клетки имеется много репликонов. В каждом репликоне можно видеть репликативную вилку – ту часть молекулы ДНК, которая под действием специальных ферментов уже расплелась. Каждая нить в вилке служит матрицей для синтеза комплементарной дочерней цепи. В ходе репликации вилка перемещается вдоль материнской молекулы, при этом расплетаются новые участки ДНК. Так как ДНК-полимеразы могут двигаться лишь в одном направлении вдоль матричных нитей, а нити ориентированы антипараллельно, то в каждой вилке одновременно ведут синтез два разных ферментативных комплекса. Причем в каждой вилке одна дочерняя (лидирующая) цепь растет непрерывно, а другая (отстающая) синтезируется отдельными фрагментами длинной в несколько нуклеотидов. Такие ферменты, названые в честь открывшего их японского ученого фрагментами Оказаки , сшиваются ДНК-лигазой, образуя непрерывную цепь. Механизм образования дочерних цепей ДНК фрагментами называют прерывистыми.
Потребность в затравке ДНК-полимераза не способна начинать синтез лидирующей цепи, ни синтез фрагментов Оказаки отстающей цепи. Она может лишь наращивать уже имеющуюся полинуклеотидную нить, последовательно присоединяя дезоксирибонуклеотиды к ее 3’-ОН концу. Откуда же берется начальный 5’-концевой участок растущей цепи ДНК? Его синтезирует на матрице ДНК особая РНК-полимераза, называемая праймазой (англ. Primer – затравка). Размер рибонуклеотидной затравки невелик (менее 20 нуклеотидов) в сравнении с размером цепи ДНК, образуемой ДНК-поимеразой. Выполнившая сво. Функци. РНК-затравка удаляется специальным ферментом, а образованная при эом брешь заделывается ДНК-полимеразой, использующей в качестве затравки 3’-ОН конец соседнего фрагмента Оказаки.
Проблема недорепликации концов линейных молекул ДНК. Удаление крайних РНК-праймеров, комплементрных 3’-концам обеих цепей линейной материнской молекулы ДНК, приводит к тому, что дочерние цепи оказываются короче 10-20 нуклеотидов. В этом и заключается проблема недорепликации концов линейных молекул.
Проблема недорепликации 3’-концов линейных молекул ДНК решается эукариотическими клетками с помощью специального фермента – теломеразы .
Теломераза является ДНК-полимеразой, достраивающей 3’-концылинейных молекул ДНК хромосом короткими повторяющимися последовательностями. Они, располагаясь друг за другом, образуют регулярную концевую структуру длинной до 10 тыс. нуклеотидов. Помимо белковой части, теломераза содержит РНК, выполняющую роль матрицы для наращивания ДНК повторами.
Схема удлинения концов молекул ДНК. Сначала происходит комплементарное связывание выступающего конца ДНК с матричным участком теломеразной РНК, затем теломераза наращивает ДНК, используя в качестве затравки ее 3’-ОН конец, а в качестве матрицы – РНК, входящую в состав фермента. Эта стадия называется элонгацией. После этого происходит транслокация, т.е. перемещение ДНК, удлиненной на один повтор, относительно фермента. Следом идет элонгация и очередная транслокация.
В результате образуются специализированные концевые структуры хромосом. Они состоят из многократно повторенных коротких последовательностей ДНК и специфических белков.


